
|
I am a biological anthropologist with an academic foundation spanning philosophy, music, physics, computers, networks, *omics, statistics, biology, and numerous social sciences. These days, I am most interested in how arboreal habitats influence primate morphology, life history, acoustical signaling, and other behaviors. My current focus is on what acoustical aspects of musical signals indicate about cognition and how certain forms of locomotion may have played a role in this co-evolutionary dynamic. |
Google Scholar: ev_vTRoAAAAJ Research Gate: David_Schruth Academia.com: DavidSchruth ORC-ID: 0000-0002-0512-450X David M Schruth on Facebook dschruth * anthropoidea * org @dranthropoid on X [Twitter] dschruth * uw * edu |
EDUCATION
with a Concentration in Statistics through CSSS
THESES
Chapters (roman numerals)
Pre-prints (lower-case)
Talks (greek)
Posters (numbers)
Software Manuals (currency symbols)
Protocols (coptic)
PUBLICATIONS
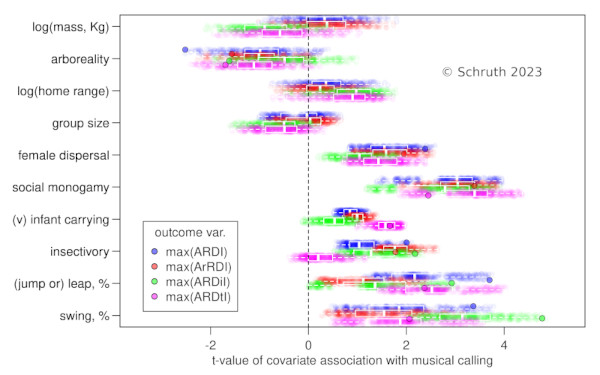
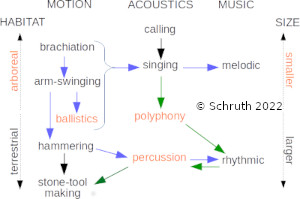
Animals communicate acoustically to report location, identity, and emotive state to conspecifics. Acoustic signals can also function as displays to potential mates and as territorial advertisement. Music and song are terms often reserved only for humans and birds, but elements of both forms of acoustic display are also found in non-human primates. While culture, bonding, and side-effects all factor into the emergence of musicality, biophysical insights into what might be signaled by specific acoustic features are less well understood. Here we probe the origins of musicality by evaluating the links between musical features (structural complexity, rhythm, interval, and tone) and a variety of potential ecological drivers of its evolution across primate species. Alongside other hypothesized causes (e.g. territoriality, sexual selection), we evaluated the hypothesis that perilous arboreal locomotion might favor musical calling in primates as a signal of capacities underlying spatio-temporal precision in motor tasks. We used musical features found in spectrographs of vocalizations of 58 primate species and corresponding measures of locomotion, diet, ranging, and mating. Leveraging phylogenetic information helped us impute missing data and control for relatedness of species while selecting amongst candidate multivariate regression models. Results indicated that rapid inter-substrate arboreal locomotion is highly correlated with several metrics of music-like signaling. Diet, alongside mate-choice and range size, emerged as factors that also correlated with complex calling patterns. These results support the hypothesis that musical calling may function as a signal, to neighbors or potential mates, of accuracy in landing on relatively narrow targets.
E: Schruth, D.M., Templeton, C.N., Holman, D.J., Smith, E.A. (2024). "The origins of musicality in the motion of primates," American Journal of Biological Anthropology. DOI: 10.1002/ajpa.24891
2023
|

|
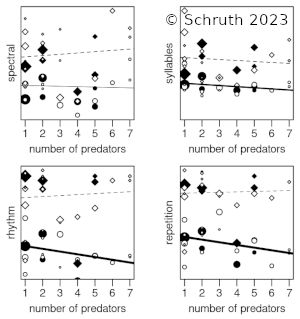
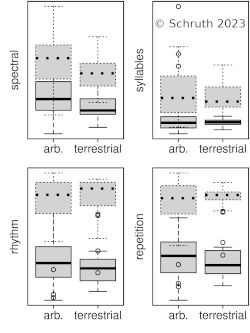
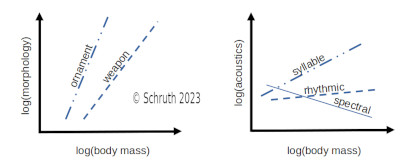
The predator-prey relationship is a fundamental directional dyad underlying trophic dynamics that emerge within food-chain networks. Numerous anti-predational behaviors are possible to help animals mitigate the risk of becoming prey, including crypsis, warding, avoidance, evasion, and confrontation. Most primates leverage both vigilance and alarm calling within groups as well as infant carrying amongst protected trees to avoid being targeted and pursued by predators. Other vocalizations of primates vary greatly and many of them contain myriad manifestations of musical complexity including spectral and temporal patterning as well as diverse contouring. Smaller primates, who may inhabit attenuated terminal branches, use more subtly short melodic sequences to maintain within-group contact, but also to circumvent eavesdropping by predators. Larger primates, who may brave open ground, use calls that are more salient, rhythmic, and syllabically diverse to assert dominance outside the group and possibly towards other species. Body size also tends to scale allometrically with other (usually morphological) traits within an individual—steeply with sexual traits and more shallowly for weaponized traits. The stronger positive correlations of body size with syllable than with rhythm suggests an attractive function of melodic elements and a mildly repulsive function for rhythm. This finding is further backed by affiliative contexts for other spectral aspects (e.g. transposition) and more threat-based contexts for temporal ones (e.g. repetition). Complex rhythms appear more often in larger terrestrial species (e.g. hominid apes and some lemurs), bolstering a possibly analogous purpose, of warding-off predators, by newly terrestrialized hominins. This combination of familial arboreality and larger-group terrestriality may have spawned our uniquely dualistic musicality—as composed of both amiable melodies and imposing rhythms.
V: Schruth, D.M. (2023). "Trophic level, arboreal position, and size of both predators and primates gave rise to a diverse and multi-functional musicality (Chapter [PDF])," In Jordania, J., & J. Wade (Eds.) Defense Strategies in Early Human Evolution, Materials of an International Conference (Full Book [PDF]), 23-26 June, 2023. Pg 169-191. Tbilisi, Logos.

|
|
|
|
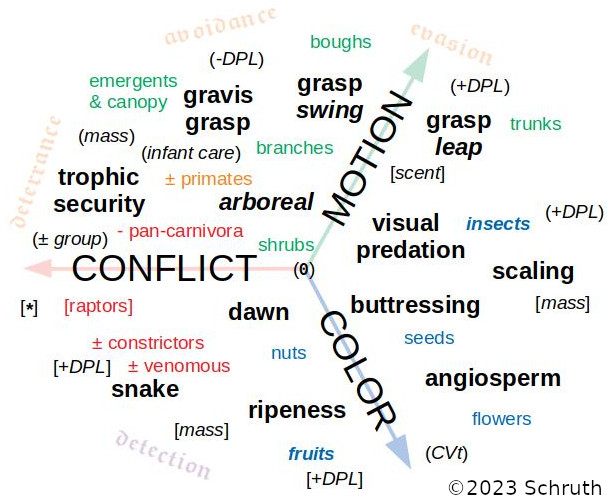
Animals create salient acoustic displays as a conflict avoidant way to secure mates or territory. Non-scentmarking anthropoids, for example, can use spectrally-rich signals to efficiently relay location, identity, condition, emotion, and intentions across distances separating callers. Arboreal primates, who are not usually the largest animals in a given geographic area, will signal from protected vantage points so as to not also attract terrestrial predators. Such a trophic security strategy can also involve nurturally protected increases in size or inter-substrate locomotion as added means of predator deterrence and avoidance. Recent work highlights acoustical associations with certain discontiguous forms of locomotion (e.g. leaping) in arboreal strategies that successfully avoid scent-stalking. I investigated sensory, habitat, morphological, ecological, and locomotor factors of primate life as driving complex acoustic display. While melodic display did associate with arboreal acrobatics, complex rhythmic display surprisingly appears more often in bipedally-capable terrestrial lineages (e.g. gorilla and some lemurs). This suggests that rhythmic complexity may emerge in exceptionally trophic-secure species with semi-vestigial limbs formerly used in more routine rhythmic locomotion. Thus, hindlimb-decoupled forelimb displays such as chest-beating in gorillas or dancing in humans could derive from a freeing-up of these accessory appendages, which support only occasional climbing, carrying, or tool use. In humans, the threat of weaponized forelimb tools may have further allowed for canine atrophy and extended terrestriality. Such stable grounding undoubtedly emboldened more aggressive hunting and audacious dancing practices in tandem with an evolution towards acoustic displays in larger groups with specialized singers and instrumental musicians.
η: Schruth, D.M. (2023). "The role of trophic security and arboreal descent in the evolution of complex acoustic display," Early Human Defense Strategies (1st bi-annual conference), Tbilisi, Georgia. [Extended Abstract (w/ figures)] DOI: 10.13140/RG.2.2.32499.30240

|

|
Primates are unusual among animals in that they have orbits that are dedicated in a forward-facing orientation. I hypothesized that various, largely family originating, forms of security from predation relaxed constraints on peripherally vigilant vision. This idea is explored as a part of the rear/oblique attack risk reduction module. I used data on orbital convergence [OC] from upwards of 88 primates and compared these values against various forms of protection including arboreality, body mass, group size, infant care, and canopy gap spanning locomotion. Body mass and infant care (e.g. carrying, weaning time) were strongly associated with higher OC across all primates, whereas group size may have enabled higher OC in smaller anthropoids and arboreality may have enabled higher OC among smaller primates. Swinging (esp. brachiating) and bridging (in Lorisidae) primates also showed higher OC values, where as leaping only associated with higher OC among larger primates. Higher values for OC among diurnal, color sensing, and angiosperm feeding primates also suggest that dietary factors may have played a role in the evolution of further OC in anthropoids. These findings are consistent with the idea that reduced predation from behind, above, and the side could have allowed primates to prioritize the target of their stereoscopic vision on items in their foreground field of view.
ι: Schruth, D.M. (2023). "Protection from predation permitted forfeit of the peripheral field and favored forward facing focus," International Primatological Society (bi-annual meeting), Kuching, Sabah, [Bornean] Malaysia. DOI: 10.13140/RG.2.2.24949.55521
In altricial species, young are born underdeveloped and mature thereafter with help from parents. Many taxa of (smaller) animals birth infants that are born helpless: marsupials, rodents, passerine birds, and most primates shelter their young through prolonged periods of immaturity. Infant carrying constitutes an unusual form of such maturation-enabling care among lower-parity terrestrial animals. In primates, ventral infant carrying, in particular, is associated with complex vocalizations and binocular vision. I hypothesized that higher risks of gravitationally accelerated impacts by (non-parachuting but) airborne euarchonta eventually led to the emergence of cognition necessary for integration of redundant sensation of both complex visual and auditory patterns. A comparison of species in our grandorder corroborates this evolutionary scenario. Unlike leaping primates who carry their infants (e.g. anthropoids), nesting non-leapers (e.g. Scandentia) and winged carriers (e.g. Dermoptera) both have minimally repetitive calls but lower orbital convergence. This suggests that both landing velocity as well as frequency may independently drive both vocal and visual evolution.
θ: Schruth, D.M. (2023). "Movement and communicative implications of ventral carrying in animals with altricial infancy," Animal Behavior Society, (annual meeting), Portland, Oregon. DOI: 10.13140/RG.2.2.35854.74569
Primates are characterized by generally orthograde postures and frequently longer hind legs (or arms) for specialized inter-branch locomotion. Cranial features of primates trend towards larger brains, a reduced snout, and convergent eyes. Primate origins hypotheses suggest such crainial configurations likely improve stereoscopic vision for high-impact limb landing. A recent musicality origins hypothesis suggests that proficiency in such binocular image resolution could be signaled by vocal production of a diversity of reappearing sound contours. I hypothesized that infant fur-cling carrying contributes to the origins of both of these (vocal and morphological) traits as it intensifies the physics involved and reflexes required for associated grasp-landing tasks. Using phylogenetically controlled regressions, I reanalyzed an amalgamation of socio-ecological datasets including data on infant carrying. To cover all primate taxa, while still accounting for the presumably more hazardous (ventral) form of carrying, I combined the results of two different studies. My formulation adds these independent ordinal measures together as: park (0), both (1), or ride (2) plus dorsal (1), mixed (2), or ventral (3), totaling to integers ranging from zero to five. Ventral carriage information was not available for non-anthropoids, but was added in post-hoc for lorises and Indri. Multivariate regression results highlight ventral infant carrying, along with mass, leaping, and swinging, as significant predictors for both outcome variables. This suggest ventral infant carrying, like other impactfully hazardous locomotion, may factor as a key influence on primate evolution. Future studies should include information on duration of carriage, weaning times, and higher resolution data for tarisers.
ζ: Schruth, D.M. (2023). "The coevolution of ventral infant carrying with primate vision and vocal signaling," American Association of Biological Anthropologists (annual meeting), Reno, Nevada.
DOI: 10.13140/RG.2.2.17819.23843
2022
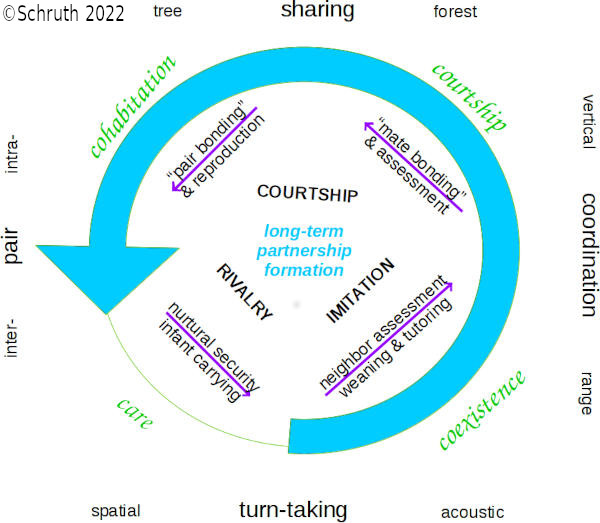
Chemical sensing via olfaction constitutes a most ancient form of inter-organism communication. But acoustical signaling via tonal and rhythmic patterning is also common among higher vertebrates. Animals that live in well ventilated habitats and move in diasporic ways have further evolved more spectrally varied and discretized call structure. But unlike song in birds, researchers have struggled to locate isolated nucleii specialized for music cognition. The brain stem, midbrain, hindbrain, and forebrain, however, all largely associate with aspects of musical performance, perception, memory, and emotion. I hypothesized that spectral features of musical display evolved as honest signals of spatial cognition for precarious locomotor tasks associated with nurturing and protecting vulnerable offspring. I investigated possible connections between motor, visual, and spatial cognitive areas in relation to both signaler production and receiver processing of acoustical features of musical output. Brain component volume fractions of 42 parts from 48 primates were compiled, from a single source, and compared against a vocal complexity index (ARDI) as well as individual musical feature scores: including tone, interval, repetition, transposition rhythm, and unique syllable count. Structures for spatial and visual perception as well as motor control and emotional processing associated moderately with areas used by species who produce calls with both temporal and spectral musical features. These findings are consistent with a dual (both receiver- and signaler- side) function of musical signals. Associations with spatio-social areas (e.g. schizocortex and insula) support direct selection for a paralimbic-based neighbor orienting [PIANO] sensory modality for mapping and anticipating movement of fellow arboreal cohabitants. Associations with motor areas (e.g. LGN, mid-brain, and thalamus) support the complementary model that signaler capacities for spatio-motive emplacement [ME] are indirectly selected by conspecific receivers. This dual manifestation in low-parity species that locomote in diasporic ways through (arboreally) diffuse habitats, is compatible with musicality serving as courtship signals by long-term mates with consistent and reliable spatial capacities directly relevant to care of vulnerable (arboreal) offspring.
i: Schruth, D.M. (2022). "Neuroecological selection for musical features through spatial reciprocity in long-term partnerships," BioRxiv (preprint). DOI: 10.1101/2022.12.13.520337
Most gibbon species produce salient duet calls at daybreak. Duets start with low frequency barks by males, followed by the female great call, and end with a short, and often complex, male-dominated coda. The female great call itself typically climaxes via a crescendoing increase in pitch, tempo, or both and characteristically features bilaterally symmetrical parabolic structures which can manifest both in the distribution of vocal units over time as well as in frequency. Male codas appear to anticipate and even complete many of these female-initiated parabolas. Employing spectrograms of species-typical great calls from nearly all gibbon species (n=12), I plotted coordinates of the upper-most frequency of each vocal unit. Using these x=time and y=frequency coordinates (plus x-differences), I tabulated the parabolas with the best possible second order polynomial fits for each species' great call. Measures of parabolic fit for each call were then compared to quantitative locomotor estimates for each species. All forms of parabolic assessment had positive correlations with leaping bout percentages across species. These results indicate that gibbon duets may function to signal a fundamental understanding of parabolic shapes--presumably useful in landing airborne locomotor (especially leaping) bouts spanning canopy elements--enabling concordant execution of arboreally projectile acrobatics.
ζ: Schruth, D.M. (2022). "Parabolic completions in gibbon duets may signal appreciation of projectile motion," Acoustical Society of America. DOI: 10.1121/10.0015580

|
|
Traits evolve in the context of historically and ecologically complex arrangements that can present difficulty for researchers attempting to uncover causal relationships or draw inference to larger populations. Primates represent a manageably diverse clade of mammals who exhibit a wide range of behaviors and morphology conducive to revealing evolutionary processes. Ideas on the origins on primates range from predation detection or deterrence to targeting in hunting or locomotion. Here, using the mmodely package on primate data for locomotion and vision, I demonstrate how the origins of primate cranial morphology can be elucidated via several ecological variables from numerous datasets. Model averaging [MA] and model selection [MS] results primarily highlight arboreal locomotor targeting and trophic security variables (such as stature or group size) as playing key roles in determining convergence of primate orbits. The package enables implementation of a combination of phylogenetic controlled regression and information theoretic (MA and MS) examination to simultaneously compare (weighted) predictor coefficients across the numerous sub-datasets generated during the exploration of all possible model combinations.
ϩ: Schruth, D.M. (2022). "Ecological factors influencing primate vision conformation: a phylogenetic regression workflow for the mmodely R-package (Version 0.2.2)," CRAN.
The historical relationships between evolved traits of organisms and the ecological settings that shape these traits are complicated systems that can be challenging to untangle. The origins of behavioral traits can particularly difficult to understand as they tend to also be mediated through the behaviors of other organisms, which are themselves constantly in flux and considerably labile. A perfect example of such a trait is that of vocal signal complexity. Animals use complex calls to assert obscured position, unique identity, special status, or emotive state to conspecifics over interference from other calls or distortions from background noise. Here, using the \Rpackage{mmodely} package on a primate vocalization dataset, I demonstrate how the origins of complex call structure, such as syllablic diversity, can be elucidated from a range of environmental and behavioral covariates from disparate datasets. Model averaging [MA] and model selection [MS] results primarily highlight locomotion and mating system as important factors driving complex calling, as well as the trophic security variables of mass, group size, and arboreality. The \Rpackage{mmodely} package enables implementation of a combination of phylogenetic controlled regression and information theoretic (MA and MS) examination to simultaneously compare (weighted) predictor coefficients across the numerous sub-datasets generated during exploration of all possible model combinations.
ϧ: Schruth, D.M. (2022). "Ecological factors influencing primate vocal signaling: a phylogenetic regression workflow for the mmodely R-package (Version 0.2.2)," CRAN.
Music is a complex acoustic phenomenon produced primarily by a single species, namely humans. However, many other species produce sounds (e.g. song-like calls and music-like note progressions) which overlap considerably with the acoustic features fundamentally present in most human societies (Darwin, 1871). A small but robust subset of key features of human music arguably includes tone, interval, rhythm, repetition, transposition, and syllabic diversity (Schruth et al., 2021). These can be found to varying degrees in the calls of non-human primates--and, when analyzed via exploratory statistical methods, have yielded a possibly zoologically applicable index of musicality useful for comparing origins of this phenomenon across species. Equipped with this newly developed acoustic (syllable) reappearance diversity index [ARDI] of musicality (Schruth et al., 2021), I endeavored to compare various levels of musical output of species with aspects of their unique ecological settings. Typical factors influencing vocalization, such as morphology (e.g. mass, age, sex, and dimorphism), immediate behavioral contexts (e.g. contact, foraging, and display), as well as more habitual ones (e.g. sex ratio, group size, monogamy, diurnality, arboreality, territoriality, predation, and acrobatic locomotion) were considered. ARDI was developed using PCA on the above-mentioned six features [see Background] prevalent in human music utterances (Schruth et al., 2021). Using ARDI scores on non-human primate vocalizations in conjunction with typical song counts in other species (e.g. whales and birds), I was able to interrogate (e.g. using multivariate linear regression) the above array of candidate ecological variables [see Aims] as evolutionary instigative. For primates, specifically, I was also able to compare brain component volumes (n=42) with vocal complexity (Schruth, 2021b).Many of the above factors were found to have a moderate influence on complex vocal signaling behavior across species. Most of these factors had positive associations with the exception of group size (mixed), sex (neutral), and predation (negative). That is, both sexes of members of small, family-sized groups (Schruth et al., 2021) that experience less predation (D. Schruth & Jordania, 2020) tend to make more musical vocalizations. Also interesting is that these species tend to live in chemically dilute environments and have reduced olfactory neural capacities. In primates the schizocortex and hippocampus have marginally positive correlations with musical (high ARDI) calls, suggesting a possible role in spatial orientation (Schruth, 2021b) as well. Most convincingly, however, visual areas (e.g. visual cortex and LGN) are even more strongly correlated with musical calling in primates, suggestive of a more locomotion relevant signaling function (Schruth, 2021b). Primarily, animals appear to have initially evolved complex vocalizations to compensate for a loss of olfaction in chemically dilute environments. This compensation may have manifested as articulation and discretization of distinct syllabic acoustic structures. Secondarily, animals appear to use musical calls to remain in acoustic contact and spatially orient with conspecifics during foraging and ranging in visually occluded environments. This second ecological pressure likely engendered highly intervalic and transpositional elements in calls to compensate for ambient noise of acoustic sound-scapes. Lastly, effective anti-predational tactics, in many vocally communicating species, could have not only required higher proficiency in evasive locomotion and associated precision in "spatio-motive landing," but also emboldened an expansion of such intra-group contact calls into more externally facing territorial displays (Schruth, 2021a). This third ecological pressure, influencing the co-evolutionary interplay of motor-eye coordination with musical-acoustic output, may have augmented levels of precision in reappearance (both repetition and transposition) for an increasingly diverse assemblage of spectral shapes.
ε: Schruth, D.M. (2022). "Adaptive origins of musical signals: olfactory compensation, arboreal orientation, and spatio-motive landing," Psychology and Music [Conference]:
Interdisciplinary Encounters. DOI: 10.13140/RG.2.2.16682.49604 You-Tube
Human-like musical behavior appears in thousands of animal species--mostly in birds but also in distant clades such as whales and primates. I investigated the idea that different aspects of human-like musicality would appear to varying degrees in non-human animals, including primates. I inspected over a thousand primate calls for evidence of acoustic features prevalent in human music. I found that syllable count, as well as transposition and repetition, were the most differentiating features between all collected calls. Furthermore, I found evidence that variation in spectrally relevant features (e.g. tone, interval, and syllable count) was likely to be more ancient than variation in more temporally relevant features (e.g. repetition and rhythm). This suggests that complex rhythmic musicality is a much more recent phenomenon--perhaps driven by group-level effects. The very first complex (non-identical unit) melodies may have been uttered by primates for tens of millions of years--initially serving as a way to accentuate individual identity. Simple (non-monotonic) rhythms--like those commonly found in the calls of most primate species--could have acted as predictable acoustic scaffolding as well as serving as a historical prerequisite for group performance. But the most complex rhythms--such as those found primarily only in human music--may serve as honest indicators of (not only) numerous participants, but groups that are able to sacrifice individualistic advertisement for the sake of larger group collective action.
26: Schruth, D.M. (2022). "Analysis of primate vocal musical complexity suggests that individual-accentuating melodies preceded group-conducive rhythms in the evolution of human music," American Association of Anthropologists. DOI: 10.13140/RG.2.2.13327.05285

|
|
|
Primates comprise an order of mammals likely originating a hundred million years ago and currently inhabit myriad environs, ranging from terrestrial to arboreal, across nearly all tropical regions of the globe. While estimates of the number of species ventures well into the hundreds, depictions of complete vocal repertoires for species are publicly available for only a fraction of these. Here I introduce such a collection of primate vocalization spectrograms amassed and standardized for future analysis. While the pool of candidate vocal studies numbers well into the hundreds, multi-spectrographic studies have been obtained for only 58 of these thus far--resulting in 1297 spectrograms for 61 species and 40 genera. Axes, labels, and any other annotations were removed and plots were organized into 842 species-specific note, phrase, and song types as determined by the primary authors. These images have been compiled into a collection of per-species tables, each including low-resolution copies of vocalization spectrograms per row, in a single PDF, each with links to the original studies. Individual spectrograms have globally unique identifiers to coordinate cross-referencing to a supplemental table detailing additional annotative information such as age, sex, and behavioral context. These data are available at a long-term data repository website [OSF.io] hosted by The Center for Open Science. This dataset uniquely enables order-wide modeling of the evolution of structural features of vocalizations in a comparative framework.
25: Schruth, D.M. (2022). "An on-line repository of spectrographic depictions of primate vocal repertoires," American Society of Primatologists. DOI: 10.13140/RG.2.2.35137.43361
Many taxa of primates produce internally and externally group-directed music-like calls. Primate infants can produce these calls--sometimes resembling the transpositional note progressions of human music. But, unlike human infants, who learn song via face-to-face visual cues, primates tend to acquire their musicality while blindly clinging to their arboreal mother's fur. Nailed primates embrace trees, branches, and fur--hindering any human-like learning. The prolonged security of infant attachment, however, may have driven music's origins, as an auditory compensator for the loss of this physical mode of anti-predation. We posit an initial musicality emerging during infant separation in such arboreal environs. Data on over a thousand primate calls, alongside ecological and musical complexity measures, confirms this thesis. When compared with long-call metrics, our melodic metric implicates mothering, locational, and exploration contexts as relevant to such internally directed calling. In conclusion, attachment serves as anti-predator security which infants manifest vocally as musically structured calls to facilitate parental orientation during initial locomotor forays.
δ: Schruth, D.M. and Dissanayake, E. (2022). "Secure arboreal and maternal attachment in primates as pre-adaptive for infant associated musicality?," Animal Behavior Conference. DOI: 10.13140/RG.2.2.33459.71202
Behavioral ecology is the science of modeling a species' adaptive fit of their behavior to their environment. Primates exhibit a vast array of behavioral modes and live in a wide variety of habitats across the world, but primarily in forested environs. Consequently, such a profusion of possible tree species exerting selection pressures on this array of primate positional and communicative modes challenges researchers with myriad habitat considerations. Presumably, such a wide variety of tree shapes, sizes, and statures should exert a corresponding diversifying selection on the behavior of its residents. This exceptional behavioral diversity of primates, incidentally makes them ideal models for testing evolutionary theories that interpenetrate the noesis of human behavior. Unlike our species, gibbons sleep in tall trees--that emerge through the forest canopies of southeast Asia--which also often serve as the primary setting for their exceptional vocal displays Many pair bonded primates participate in coinciding vocal behavior that often manifests in the form of calls with interacting male and female contributions. Socially monogamous gibbons, tarsiers, and callitrichids produce mutable vocal duets that feature such acoustic patterning. Gibbons, in particular, routinely exhibit duetting behaviors in all but a few species. The females' "great call" forms the center-piece of such elaborate displays--often featuring a diversity of syllables which typically increase in frequency and accelerate into a rapid series of upward frequency sweeps blurring repetition into both transposition and trill. Theories on the function of these calls range from resource spacing, to pair-bonding and mate attraction. But few studies to date have looked at various features, especially in combination, and how display structure could relate to ecology. Such structured patterning (e.g. rhythm) of collaborative primate calls may facilitate turn-taking between mated pairs. Such joint vocal coordination could signal corresponding locomotor coordination to neighboring conspecifics. Species that duet with such acoustic features (e.g. rhythmic tempo) likely evolved mental capacities conducive to riskier arboreal locomotion in taller trees. I tested this idea with a species-level dataset of spectrographic vocal duets of gibbons--specifically, assessments of musical features (e.g. repetition, transposition, and syllable) present in each song. For each gibbon species, these features were compared with a dataset of associated indigenous species of "emergent" trees from genus Shorea (of family Dipterocarpaceae) Surprisingly, only a few of the features that distinguish primate calls as musical (e.g. rhythm and syllabic diversity) corresponded to these ecological features--primarily only diversity and height of associated emergent tree species. Specifically, emergent height correlated the most consistently with the spectral complexity of the protracted (female) components of the gibbon great call. Contrary to my main prediction, however, arboreal height did not correlate positively with musical rhythm, although it did with percentage of non-overlapping units. And there are marginal indications that it could instead correlate with the diversity of these gibbon-associated emergent tree species.Here, I have questioned the primacy of rhythm as the sole indicator acrobatic primates (here gibbons) use in their acoustic coordination displays (e.g. the great call duet), and a solid case is emerging for the importance of spectral features. Syllabic diversity, for example, was not ruled out as compensating for reduced chemical signaling. As humans, we constitute the only musical ground-primate and we owe our profuse variety of modern musical forms not only to cultural transmission within and between large groups, but also, ultimately, to sensory constraints inherited from our ancient olfactory-impoverished primate progenitors. The additional, and somewhat serendipitous finding--that arboreal (emergent) variety and height correlates with musical complexity in the longer female-component of gibbon great calls--provides additional support for an arboreal locomotion conducive cognition signaling related function of primate duets.
γ: Schruth, D.M. (2022). "Primate duet display via arboreal locomotor predictability: emergent height and variety as selecting for more complex gibbon great calls," Conference on Interdisciplinary Musicology. DOI: 10.13140/RG.2.2.20037.93921
Ecological investigation is invariably plagued by the inherent difficulty of collecting and analysing all possibly relevant causal input variables. While many datasets have been amassed to answer ancient ecological mysteries, and numerous tools have been developed to enable merging of such disparate sources of possible influence--these problems remain somewhat unresolved due to inaccessibility to tools and data. For example, current understanding on when primates originated is uncertain and the extent to which possible ecological influences (e.g. leaping, daylight, canopy height, or diet) most influenced the their emergence, is likewise unclear. A database that has recently become available online derives from the "All the Worlds' Primates" project. This database, however, is developed in SQL and currently requires the use of a web interface with limited functionality. Here I showcase an additional tool, the "primates" r-package which enables easy-to-use SQL-style functional access to these data. To highlight it's utility, I review several primate origins puzzles, review some possible solutions, and demonstrate how the package functionality helps bring together disparate datasets compiled by thousands of independent specialized primatologists--to help begin to answer these age old questions with big data all via a familiar R interface.
24: Schruth, D.M. (2022). "The 'primate' R package enables easy merging of All the Worlds Primates via SQL tables.," Use-R Conference. DOI: 10.13140/RG.2.2.21715.66080
2021
Music is especially valued in human societies, but music-like behavior in the form of song also occurs in a variety of other animal groups including primates. The calling of our primate ancestors may well have evolved into the music of modern humans via multiple selective scenarios. But efforts to uncover these influences have been hindered by the challenge of precisely defining musical behavior in a way that could be more generally applied across species. We propose an acoustic focused reconsideration of "musicality" that could help enable independent inquiry into potential ecological pressures on the evolutionary emergence of such behavior. Using published spectrographic images (n=832 vocalizations) from the primate vocalization literature, we developed a quantitative formulation that could be used to help recognize signatures of human-like musicality in the acoustic displays of other species. We visually scored each spectrogram along six structural features from human music--tone, interval, transposition, repetition, rhythm, and syllabic variation--and reduced this multivariate assessment into a concise measure of musical patterning, as informed by principal components analysis. The resulting acoustic reappearance diversity index (ARDI) estimates the number of different reappearing syllables within a call type. ARDI is in concordance with traditional measures of bird song complexity yet more readily identifies shorter, more subtly melodic primate vocalizations. We demonstrate the potential utility of this index by using it to corroborate several origins scenarios. When comparing ARDI scores with ecological features, our data suggest that vocalizations with diversely reappearing elements have a pronounced association with both social and environmental factors. Musical calls were moderately associated with wooded habitats and arboreal foraging, providing partial support for the acoustic adaptation hypothesis. But musical calling was most strongly associated with social monogamy, suggestive of selection for constituents of small family-sized groups by neighboring conspecifics. In sum, ARDI helps construe musical behavior along a continuum, accommodates non-human musicality, and enables gradualistic co-evolutionary paths between primate taxa--ranging from the more inhibited locational calls of archaic primates to the more exhibitional displays of modern apes.
D: Schruth, D.M., Templeton, C.N., and Holman, D.J. (2021). "On reappearance and complexity in musical calling," PLoS ONE. DOI: 10.1371/journal.pone.0218006

Primate vision is thought to have evolved in connection with life in the trees. However, several inter-related origins theories--those addressing possible co-evolution with size, predation, diet, daylight, locomotion, and groups--also provide reasonable explanations of their distinct cranial-visual morphology. We hypothesized that demand for high-speed arboreal grasp-landing facilitated predation avoidance thereby reducing the need for lateral facing orbits. To test this proposed influence, in the context of a likely very multi-causal adaptive landscape, we consolidated published data on extant primate species including body mass, daily path length, arboreality, insectivory, frugivory, activity period, leaping, swinging, and group size. Phylogenetically controlled regressions, on three different taxonomic subsets of the primate order, highlight size and environmental influences as the most compelling factors explaining higher orbital convergence [OC]. Moreover, activity period and group size (in anthropoids) as well as arboreality and body mass (in non-anthropoids) associated convincingly with higher OC. After considering size and path length, suspensory (and to a lesser extent leaping-based) grasp-landed locomotion co-varied with OC, primarily in anthropoids. Nocturnality had negative, and leaping mixed, associations with OC--thus, with the exception of those relating to nocturnal-locomotion, all adaptive origins theories considered were at least partially corroborated. The conflicting associations of OC with leaping, is largely attributable to the exceptionally small (and more taxonomically contentious) members of the order. But prospects otherwise remain bright for our two freshly illuminating theories of grasp-swing, in anthropoids, and rear attack risk reduction [RARR], more generally, as they provide compelling alternatives to sized based models (e.g. predation deterrence and allometric scaling) in explaining deep divisions in the primate order.
h: Schruth, D.M. (2021). "Arboreal locomotion and trophic security at the dawn of Euprimate vision," EcoEvoRxiv. DOI: 10.32942/osf.io/d6wk2
This protocol provides a method to realize phylogenetic control in multivariate regression modeling while estimating tree transformation parameters en route. The protocol requires compiling a list of all possible variable combinations (at all model lengths) and iterating through these while estimating the transformation parameters along side the regression. A combination of AIC and the coefficient of determination can be used to select the "best" model from numerous possible model lengths. The average of the tree transformation parameters can then be used on these "best" models to perform the final phylogenetically controlled multivariate regression.
ϥ: Schruth, D.M. (2021). "A global variable-permutation based approach for estimating tree transformation parameters used in phylogenetically controlled multivariate regression," Protocols.io. DOI: 10.17504/protocols.io.bzdhp236
Most vertebrate animals are adept at passing air over constricted nasal or throat passages to produce acoustic vibrations in order to communicate position and identity to conspecifics. More complex, and perhaps music-like, patterning in the form of displays might also possibly function as honest signals of underlying cognition. Arboreal primates and birds, as well as aquatic mammals--including dolphins, whales, seals, penguins--make such calls. Increasingly researchers are honing in on specific aspects (e.g. tone, rhythm, transposition, repetition) of these calls to determine when how and why they have emerged. Yet poorly understood, still, is how specific features of each of these animals' neural circuits are involved in the processing of each of these features. I hypothesized that animals should possess similarly high volume, encephalization, or utilization of brain structures (e.g. archicortex, mid-brain, and hind brain) for pattern matching of melodic content considering similarities in higher dimensional modes of locomotion. I calculated such brain component volumetric percentages of a small sample of non-human primates to investigate possible connections with spatial capacities. Using a musicality index, measuring unit reappearance diversity, both unveils the visual cortex (p<0.1), including LGN (p<0.01), and confirms spatial, hippocampus and brainstem (p>0.3), areas as positively associated with musical calling. If Tarsius is removed as an outlier, results alternatively suggest correlations with higher brain structures. For syllable count, there were significant (p<0.1) associations--positive with neocortex and negative with three different structures involved in olfaction. In conclusion, it appears that brain structures for vocal-auditory (and perhaps visual) sensory integration may have out-prioritized structures for olfactory communication for space in the evolving primate brain. The subtle tarsier-inclusive based results are consistent with the idea that music-like signaling evolved with fundamental audio-visual, motor control, and orientating structures of the midbrain and allocortex in more ancient primate linages.
23: Schruth, D.M. (2021). "Primates evolved spectrally complex calls in compensation for reduction in olfactory cognition," Proceedings of the Annual Meeting of the Cognitive Science Society, 43. Retrieved from https://escholarship.org/uc/item/0jw446s9
[poster (PDF)]
Musical behavior appears across the animal kingdom in organisms as diverse as insects, birds, and many mammals including primates (Clark 1879; Darwin 1871; Marler 2000). While a minimal amount of sociality, at least a sender and a receiver, is indubitably prerequisite in acoustic signaling systems (Johnstone 1997), ultimate ecological causes for musical behavior are still poorly understood. The development of quantitative methods of measuring musical behavior--including intervalic, rhythmic, and melodic content of calls--is also still in its infancy (Schruth, Templeton, and Holman 2019). Here we primarily measure more melodic and song-like aspects of calling behavior by looking at how similar vocal units connect to form a diversity of clustered acoustic types.I hypothesized that habitat based influences--such as sucessful substrate connectivity during precarious and high-speed arboreal locomotion--would correlate more strongly with musical behavior measured using these more melodic musicality metrics. Along the way, I also hoped to understand the strengths and weaknesses of several different vocal complexity measures--which capture melodic aspects of singing behavior--in order to elucidate which features most closely associate with certain contexts.Here, using vocalizations of 57 species of primates and their ecological settings, I compare several indexes of vocal complexity. These include the newly developed acoustic reappearance diversity index [ARDI] emphasizing redundancy, a euclidean distance based song complexity index [SCI] emphasizing call length (Sawant et al. 2019), and the simplest complexity measure of unique syllable count. ARDI is calculated by the (PCA reduction determined) structural acoustic universal human musical features of repetition [r], transposition [t], and syllable [s]--formulated as (r+t)*s (Schruth, Templeton, and Holman 2019). SCI can be approximated using only syllable count [s] and total length [L] via s*sqrt(L^2 - s*((L/s)-1)^2)). Syllable count is simply the sum total of spectrally unique unit-shapes within a call (Templeton, Laland, and Boogert 2014). I reaffirm validation of this acoustic diversity measure, ARDI, as corroborating call names and types (e.g. "musical" or "song") ascertained by the primary researchers, as matching well with species previous judged as being musical, and by being compellingly correlated with more traditionally-geared bird-song complexity measures. Differences include SCI's tendency to highlight more repetitive calls versus ARDI's tendency to highlight shorter, more transpositionally melodic calls. I also distinguish them by noting that SCI more deeply discounts chorusing primates in comparison to both ARDI and syllable count alone. All metrics confirm earlier observations that social monogamy is decidedly the most strongly associated with music-like behavior. But only ARDI is strongly differentiating of habitat effects such as forest composition. Furthermore, ARDI seems to associate most strongly with display calls produced by females.I confirm the idea that a multitude of selection pressures, including both the acoustics of wooded habitats as well as parental and other within-deme social influences--including modest socially-monogamous conjoinment factors--likely drove the evolution of melodic behavior in primates. In particular, ARDI indicates that primate musicality could best be exemplified by female-centric calls from socially monogamous groups of small size. ARDI, in comparison to SCI, also tends to differentially highlight arboreal and particularly wooded contexts. I extrapolate upon these findings by suggesting that short, transpositionally music-like (ARDI leaning) calls were quite ancient, perhaps resembling those of modern galagos, and could have preceded the longer more song-like (SCI leaning) calls in later, derived linages of calling primates (e.g. indri, gibbon, and tarsier). In counter-point, however, our markedly extreme human emphasis on transpositionally noted scales could imply that the more repetitive (and presumably gibbon-like) singing bouts of our ancestors may currently be under less positive selection in our species.
22: Schruth, D.M. (2021). "Diversity in connectivity of primate vocal units: associations with socio-ecological contexts," Proceedings of ICMPC-ESCOM, University of Sheffield, UK.
Over a hundred million years ago, primate predecessors may have been geographically distributed throughout both the Gondwanan and Laurasian supercontinents. Gondwana included both the future land masses of India and Madagascar whereas Laurasia spawned many islands of the Malay archipelago, including Java, Borneo, and Sulawesi. Under all radiative scenarios, singing primates appear to have evolved (and survived) on these smaller and sub-continental islands adjoining the Tethys (now Indian) ocean. Predation is hypothesized to play a role in selecting for primate species that are staturally comfortable enough to vocalize saliently. And protective effects, such as arboreality and size, may therefore indirectly play a predation deterring role--as evidenced by the example of Indri, the largest mammal on the subcontinent. In the Malay archipelago, however, even the smallest of primates, tarsiers, are frequent vocalizers--perhaps resulting from the loss of anti-predator silencing due to protective isolation on small islands lying largely east of the Wallace and Huxley Lines. Furthermore, a loss of sense of smell that tends to accompany discontiguous locomotion is also associated with changes towards more discretized and higher syllable count vocalizations. Indri, tarsiers, gibbons, and galagos all produce discrete syllables that reappear both temporally and spectrally, corresponding to repetition and transposition, respectively. Vocal-motor control for such calls could plausibly serve as signals of analogous spatio-motor capabilities useful for visual focus on landing targets such as branches. Along these initial lines of inquiry, I synthesized the results of numerous related studies to more fully circumscribe a zoologically-broad and multi-factorial theory on the origins of musical calling. In addition to stressing the important considerations of predation and olfaction, I conclude by suggesting that the more general neologism of "motive landing" could apply more inclusively to other non-primate and non-avian surface-landing tetrapods such as cetaceans, pinipeds, and sphenisciformes who also produce such calls. I end by exploring how the descent of man from trees may have freed hominin hands for percussion--enabling creation of uniquely human music that is both more rhythmically complex, lower in frequency, and louder in amplitude.
β: Schruth, D.M. (2021). "Musical calling as a signal of motive landing ability in diasporic tetrapods inhabiting upper trophic levels," Evolution, a joint meeting of the American Society of Naturalists, the Society for Systematic Biology, and the Society for the Study of Evolution. (Virtual Conference). DOI: 10.13140/RG.2.2.29378.25280/1
[
talk (MP4);
transcript (PDF)
]
Many animals produce sounds with song-like qualities to communicate who and where they are to conspecifics when their senses of smell, and vision, are rendered ineffectual across long, occluded distances. Social factors drive song evolution, but we consider the neglected effects of habitat which are difficult to observe, measure, and analyze. In particular, predation, trophic rank, and size inequalities could manifest as key evolutionary influences in determining motivations for vocalizing as doing so can sacrifice the position of callers. We explore how singing cetaceans, birds, and primates avoid predators using size, flight, and trees. We also posit a replacement of arboreal protection with ballistics and group intimidation as the key shift in human evolution towards protomusical rhythm. Diet, broadly expanded to also include both predation risks and payoffs, could be construed as an overarching evolutionary force driving the evolution of singing and protomusical behavior.
21: Schruth, D.M., Jordania, J. (2021). "Extreme predation reduction as a selection effect for singing behavior," Animal Behaivor (twitter conference). [tweets (PDF)]
21: Schruth, D.M., Jordania, J. (2021). "Extreme predation reduction as a selection effect for singing behavior," Animal Behaivor (twitter conference). [tweets (PDF)]
2020
Music is a singular behavior humans exhibit via the creation of patterned sounds organized, for example, along rhythmic and melodic dimensions. Music-like behavior, however, also appears in many other animals including non-human primates. Calls with western musical attributes are primarily ascribed to species of gibbon, tarsier, indri, and potentially also marmoset, tamarin, and titi monkeys. A recent survey of primates, using a continuous measure of acoustic musicality, suggests that many other species--including galagos, several lemurs, and some old-world monkeys--also exhibit calls with music-like qualities. Based on this ubiquity and a recent arboreal-origins theory for musical behavior, I hypothesized that the ancestor of all living primates was likewise also somewhat musical. Ancestral character estimation was used to reconstruct ancestral phylogenetic states using scored spectrographic depictions of vocalizations of extant primates (n=58 species) to determine if musical calling is the initial primate condition. The results suggest that nearly all primate families have an ancestor with at least one moderately music-like call--two or more reappearing syllables--with the clear exception of Cercopithecine. These cheek-pouch monkeys likely experienced a multi-million year diminution in musical behavior, perhaps in conjunction with their recent adaptations for quadrupedal terrestriality. These findings largely indicate that exhibition of at least one music-like call is a normal, and likely quite ancient, constituent of primate vocalization behavior.
g: Schruth, D.M. (2020). "Musical calling as a behavior ancestral to all modern primates ," PsyArXiv.
DOI: 10.31234/osf.io/mkze8
DOI: 10.31234/osf.io/mkze8

A surprisingly diverse array of animals produce sounds with song-like qualities in order to communicate who and where they are to conspecifics when their senses of smell, and in some cases vision, are rendered ineffectual across long or occluded distances. Social factors are typically considered to drive the evolution of such calls, but here we consider broader effects of habitat which are often difficult to observe, measure, and analyze, and are thus typically neglected. We tested the hypothesis that ecological protections from predation enabled animals to call by neutralizing the risk inherent in vocally forfeiting position. We compiled data on human and non-human singing species that live in arboreal, aerial, or aquatic (i.e. non-planar) habitats, to measure the degree to which each presumably helps to reduce predation and thereby allow singing. We find that singing species also forfend predation via larger size as well as increased levels of arboreality and flight. Arboreal primates and birds do appear to sing significantly more so than terrestrial ones. Hunting and warfare in humans, respectively, likewise correlate with the musical features of rhythmic and melodic tension. In all species explored, however, only body size universally correlates positively with singing behavior. We conclude that effects of predation, especially when viewed in a larger, food-web context, play a central role in determining who sings and who does not.
f: Schruth, D.M., Jordania, J. (2020). "Singing behavior via reduced predation risk," PsyArXiv.
DOI: 10.31234/osf.io/u9m8z
DOI: 10.31234/osf.io/u9m8z
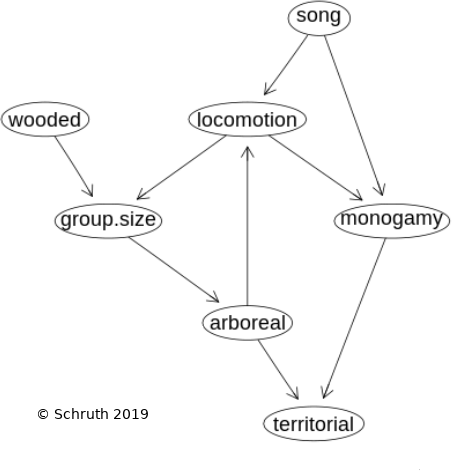
Animals communicate acoustically to report location and identity to conspecifics. More complex patterning of calls can also function as displays to potential mates and as territorial advertisement. Music and song are terms often reserved only for humans and birds, but elements of both forms of acoustic display are also found in non-human primates. While theories on proximate functions abound, ultimate drivers of specific call structures are less well understood. We hypothesized that spatio-temporal precision in landing during perilous arboreal locomotion favored the evolution of musical calling in early primates--vastly preceding the origin of more music-like behavior in hominoids and subsequent emergence of music in later hominids. We test this locomotion based hypothesis on the origins of proto-musicality using spectrographic depictions of vocal repertoires of modern day primates and corresponding estimates of locomotor activity. Phylogenetically controlled regression analysis of 54 primate species reveals that arboreal locomotion and monogamy are robust influences on complex calling patterns while controlling for other socioecological variables. Given that these findings rest primarily upon a handful of deep branching points in the primate tree, we conclude that this coevolution likely occurred very slowly, occupying on the order of tens of millions of years
e: Schruth, D.M., Templeton, C.N., Holman, D.J. and Smith, E.A. (2020). "Evolution of primate protomusicality via locomotion," BioRxiv. DOI: 10.1101/2020.12.29.424766
Examination into hypothetical origins of primate morphology and behavior can be challenged by many possible confounding ecological factors. Statistical modeling with multivariate and phylogenetic control can be a fruitful approach if tools and data are readily accessible. Human readable, webpage-style data sources are useful, but may not be machine readable or amenable to usage in statistical software programs. Recent systematic efforts in tabulating data on primates exist in the form of individual silos of smaller, single-table compilations or via the All The World's Primates [AWP] larger relational SQL database. While much effort has gone into organizing and curating such datasets, the downloading of data on multiple-species can be difficult or infeasible without specific technical training. With the increasing popularity of statistical analysis environments, we can bridge this gap between these modern analysis tools and the machine scalable data archives which are often coded in non-human readable ways. We introduce here a package for use in the R statistical programing environment containing tools for accessing tabular extractions from AWP along with aggregated summary datasets and demonstrations of common analysis routines. As an example, we illustrate how the pervasive hazard of arboreal fall-risk in primates is relatively neglected in studies because the data (e.g. canopy height utilization) is often sparse or absent. We further demonstrate recoding routines to transform variables (e.g. diet) into more efficiently recoded equivalents.
20: Schruth, D.M., Marc, M., and Rowe, N. (2020). "The fall-out and fruits of big-data in primate research: disentangling evolutionary causes and effects," American Society of Primatologists (annual meeting).
Unlike (litter-birthing) tree shrews, who leave their infants in nests, most (lower-parity) primates carry their infants to avoid predation. Additional avoidance strategies such as inter-tree leaping can create communication gaps--which not only allows primates to avoid scent-tracking by predators but also requires them to develop contact calls that are distinguishable from background noise. The function of acoustic differentiability goes beyond just group confirmation of location and identity, however, as extra-group display can also evoke elaborate acoustic patterning. To help disentangle these selective influences, I explored a collection of 502 calls and contexts, from 46 species, in a causal modeling framework using the PC-algorithm. I found that contact and foraging contexts had more tonal and repetitive elements while display and territorial contexts were more syllabically diverse. At the species level, diversely reappearing syllabic displays were directly associated with social monogamy and risky locomotion (e.g. leaping and swinging)--indicative of a possible three-way co-evolution. These results are consistent with the theory that parental care in primates could have facilitated the learning of precarious arboreal subsistence strategies that further necessitated highly patterned acoustic signals. While galagos and tarsiers, who park their infants, serve as important exceptions, anthropoids have been synapomorphic in fur-cling carrying for most of the Cenozoic and serve as a plausible parental attachment model underlying the co-evolution between complex limb landing and elaborate note landing in our hominoid ancestors.
19: Schruth, D.M. (2020). "Did infant-laden leaping lead to lullaby, melody, and prosody?," American Association of Physical Anthropology. Vol. 171, (S69). DOI: 10.1002/ajpa.24023
ϣ: Schruth, D.M. (2020). "Protocol for scoring animal calls on structural acoustic features prevalent in human music," Protocols.io. DOI: 10.17504/protocols.io.bp5emq3e
2019
|
This dissertation explores the social, behavioral, and habitat based influences on the evolution of elaborate vocalizations in primates. The goals of this project were to merge and test hypotheses derived from both the primate morphological origins literature and the musical behavior origins literature. Theoretical elements from the leaping and arboreal hypotheses from the primate origins literature were brought to bear on vocal display data from extant primates and on theories on the function of song in animals and music in humans. Specifically, I used computational statistical techniques to test the dual hypotheses that precision limb landing, as required in navigating complex arboreal topologies, could have not only shaped primate morphology, but also evolutionarily shaped their vocal signaling behaviors. I also tested other theories from the literature on both animal communication (e.g. the acoustic adaptation hypothesis) and human music (e.g. coalition signaling as well as group and pair bonding). I collected behavioral data, for as many species as possible, from the primary primate behavioral literature, including vocalization, locomotion, and socio-ecological regression control variables. Data were then analyzed in three separate substantive chapters. First, in chapter 2, spectrographic vocalizations were scored manually along ethnomusicologically universal acoustic parameters and subsequently reduced into a univariate acoustic reappearance diversity index [ARDI], reflecting call elaborateness. Second, in chapter 3, quantitative locomotion activity data (e.g. leaping and swinging percentages) were harvested from the positional behavior primary literature in order to both estimate its ancestral influence on morphology as well as to create better (non-binary) predictor variables for testing vocal display origin theories. Third, in chapter 4, I tested for my hypothesized co-evolutionary relationship while controlling for phylogeny. ARDI was highest in smaller, socially monogamous, and forest living groups, as well as positively associated with swinging and leaping. These results are consistent with the dual hypothesis that both elaborate vocal displays and vision changes in primates could have evolved as a result of increased demand for precision limb landing locomotion. In chapter 5, I discuss the limitations and drawbacks as well as the theoretical significance, scientific relevance, broader implications, and ideas for future research.
IV: Schruth, D.M. (2019). "Ecological Selection Pressures for Singing Behavior in Primates," Doctoral Thesis, University of Washington. Chair: D.J. Holman
Musical behavior is likely as old as our species with song-like calls possibly originating
tens of millions of years ago when primates were first radiating across the globe. Early singing
likely evolved into the music of modern humans via multiple selective events, but efforts to
disentangle these influences have been stifled by challenges to precisely define this behavior in
a broadly applicable way. Detailed here is a method to quantify the elaborateness of acoustic
displays using published spectrograms (n=832 calls) from the literature on primate
vocalizations. Each spectrogram was scored by five trained analysts via visual assessments
along six musically relevant acoustic parameters: tone, interval, transposition, repetition,
rhythm, and syllabic variation. Principal Components Analysis (PCA) was used to reduce this
multivariate assessment into a simplified measure of musical elaborateness. The resulting
"acoustic reappearance diversity index" [ARDI] simultaneously captures syllabic variation and
spectral/temporal redundancy in a single continuous variable. The potential utility of this index
is demonstrated by applying it to several social and habitat-based theories of acoustic display
origins. Our results confirm that primate species living in small, monogamous groups have
song-like calls, while forest habitat had a less pronounced association.
Ch 2 - The acoustic reappearance diversity index: from primate song to human music
Arboreal living is thought to have driven the evolution of vision improvements associated with
the origins of the primate order from the late Cretaceous up through the Eocene. We explore the
possibility that rapid, high-risk, arboreal grasp-landed locomotion could be both an initial and
continued force in euprimate cranio-visual evolution. Quantitative observations of locomotor behavior
on 130 extant primate species was used to both reconstruct locomotor activity budgets of ancestral
primates as well as to better understand the evolution of a most salient feature of the euprimate
cranium, anteriorly convergent ocular apertures. The phylogenetic mean of locomotor mode
frequencies was used to estimate ancient locomotor patterns. Fractional estimates of two rapid and
high-risk locomotor mode spectrum behaviors were "leaping" at nearly one-third (for basal euprimates)
and "swinging" at one-fourth (for basal hominoids). Although path length and diet were less significant,
phylogenetically controlled regressions highlight color vision ahead of size, locomotion, and activity
period as substantively associated factors possibly driving the evolution of orbital convergence [OC].
Thus, OC appears to serve a multiplicity of purposes, as evidenced by the result that OC was
simultaneously positively correlated with the orthogonal traits of group size and color vision. It is
possible that the many plausible, competing influences on anterior OC were separately realized because
of a relaxed requirement for lateral facing orbits--perhaps due to overall predation reduction. Less
speculatively, these results support a gradual and sequential evolutionary accretion of the key defining
morphological traits of the primate order (grasping, leaping legs, and OC) as driven by temporally
disparate ecological shifts.
Ch 3 - Precision limb landing locomotion and the origins of primate morphological traits
The complex ecological niche of arboreality, via associated locomotion through it, may have
provided the selective context for spatial-cognitive evolution in primates. We suggest that elaborate
vocal displays (i.e. singing) subsequently evolved to signal these underlying spatial proficiencies. We
test this using data on vocal elaborateness and forms of spatially demanding locomotion from 51 non-
human primate species. Vocal elaborateness was assessed by a recently developed "acoustic
reappearance diversity index" and for locomotion we used leaping and swinging frequencies. We were
able to demonstrate a positive association between aerial spectrum locomotion, such as brachiation and
jumping and leaping, with elaborate vocalizations, such as songs, duets, and great calls. We used
phylogenetically controlled regression analysis on a merged dataset which included these measures of
both vocalization and locomotion as well as other socio-ecological controls such as social monogamy
and habitat. While we reconfirm that monogamy had a significant positive association with musical
behavior in primates, forest dwelling (a proxy for habitat acoustics) was not significant after controlling
for shared ancestry. The results fail to reject our hypothesis that elaborate vocal displays could have
co-evolved with spatially demanding locomotion as signals of spatial abilities.
Ch 4 - Co-evolution of elaborate vocal display and precision landing locomotion
Darwin considered music to be the most mysterious behavior with which we are endowed and
the best example of sexual selection in humans (Darwin, 1871). This dissertation attempted to help
solve this mystery by applying principles of behavioral ecology and signaling theory to a broad base of
data and theories developed in ethnomusicology and biomusicology, as well as from the primate
locomotion, vocalization, and origins literature. I have provided a new and compelling theory in the
form of the spatio-motor signaling hypothesis presented in the previous chapter. This theory posits that
spatio-visual cognitive evolution in primates was largely driven by an increasing requirement for active
locomotion, especially in the form of leaping, between the terminal branches of floral entities. These
elaborate vocal communication systems likely evolved, initially, to both stay in contact and assert
identity across these distances, and, eventually, to signal the spatially discriminating cognition and
ocular sensory-motor proficiencies required to land such leaps.
Ch 5 - Acoustic universals, discrepant definition, and ocular-motor signaling origins
|

|
Singing has evolved independently in several different taxa of vertebrates. In primates, the ultimate co-evolutionary influences on song evolution include social signaling and habitat acoustic effects. Proximate mechanisms such as how these songs are learned, however, are less well understood. Vocal learning species include birds (songbirds, parrots, and hummingbirds) and mammals (bats, elephants, seals/sea lions, whales/dolphins, and humans). Primates, however, are not typically considered vocal/song learners. Here we test this notion by comparing adult with non-adult calls sampled from 59 primate species' spectrographic vocal repertoires (n=823) scored according to a recently developed song index. Adult calls had significantly higher diversity of reappearing syllables than non-adults (p=0.025). These results suggest that we should reconsider the established null hypothesis that the structure of adult primate songs are innate.
18: Schruth, D.M., Templeton, C.T. (2019). "Human music universals derived index suggests adult primates learn their songs," American Society of Primatologists Annual Meeting, American Journal of Primatology, Vol 82(S1).
Musical behavioris likely as old as our species with song originating as early as 60 million years ago in the primate order. Early singing likely evolved into the music of modern humans via multiple selective events, but efforts to disentangle these influences have been stifled by challenges to precisely define this behavior in a broadly applicable way. Detailed here is a method to quantify the elaborateness of acoustic displays using published spectrograms (n=832 calls) culled from the literature on primate vocalizations. Each spectrogram was scored by five trained analysts via visual assessments along six musically relevant acoustic parameters: tone, interval, transposition, repetition, rhythm, and syllabic variation. Principal Components Analysis (PCA) was used to reduce this multivariate assessment into a simplified measure of musical elaborateness. The resulting " acoustic reappearance diversity" index simultaneously captures syllabic variation and spectral/temporal redundancy in a single continuous variable. The potential utility of this index is demonstrated by applying it to several social and habitat-based theories of acoustic display origins. Our results confirm that primate species living in small, monogamous groups have song-like calls, while forest habitat had a less pronounced association.
d: Schruth, D.M., Templeton, C.T., and Holman, D.J. (2019). "A definition of song from human music universals observed in primate vocalizations," BioRxiv. DOI: 10.1101/649459
|
17: Schruth, D.M. (2019). "Evolutionary selection for song: disentangling content from context," CSSS
(departmental poster session). |
|
 |
Musical behavior predates the emergence of Homo sapiens and primitive song could have originally evolved in our order 65 MYA, convergent with and independent of its emergence in Aves and Cetaceans. The avian display call literature uses quantitative metrics such as trill rate, consistency, repertoire size, song bout length, and complexity to evaluate songs. There have been few analogous efforts to quantify aesthetic qualities of display calls in our own order. We developed a novel method to quantify the elaborateness of acoustic displays using published spectrograms of primate vocalizations (n=832 calls). These spectrograms (plots of acoustic energy across both frequency and time) were visually scored along the following ethnomusicologically universal acoustic parameters: tone, interval, transposition, repetition, rhythm, and syllabic diversity. Principal components analysis was used to reduce the scores to a single univariate measure of musical elaborateness. The resulting index is mathematically defined as the expected number of syllables reappearing within a call. Specifically it is the count of unique syllables multiplied by the probability that any given unit reoccurs over time or at different frequencies. This "reappearance diversity" index was verified as correlating well with display contexts. The utility of the index was demonstrated by testing it against popular co-evolutionary theories on the function of song such as group and pair level signaling as well as the acoustic [habitat] adaptation hypothesis. Species who group into monogamous family units (2-5 individuals) had higher index values. We found no direct support for the acoustic adaptation hypothesis.
15: Schruth, D.M., Templeton, C.T., and Holman, D.J. (2019). "Reappearance diversity: quantifying musicality in primate vocalizations," American Association of Physical Anthropologists (annual meeting), American Association of Physical Anthropologists, Vol 168(S68).
2018

2017-2016
2015
Terrestrial predation on much of the planet's two-dimensional surface can be exemplified by larger predators pursuing smaller prey who use peripheral vision along with numerous, sudden directional changes during potentially prolonged pursuit. These dynamics were dimensionally altered when primates evolved anatomy specialized for arboreal avoidance. Appendages that embrace (rather than claw into) substrate enable refuge from larger terrestrial specialists because a high body to branch size ratio can render predation infeasible in a fractally slendering terminal branch topology. An extinct family of non-burrowing, legless, large-gaped, constricting reptiles (madtsoiids) could have overcome this size ratio barrier in the late Cretaceous when grasping Euprimateformes were first emerging. Frequent leaping has been suggested as serving as an anti-predation locomotor strategy in prosimians. I extend this idea by hypothesizing that leaping specializations in primates first evolved as a snake predation avoidance mechanism. Absent substantial (terrestrial) predation from their posterior, (strictly arboreal) primates may have instead evolved anteriorly convergent vision for landing leaps as part of expeditious evasion from this new serpentine threat to its terminal branch refuge. I test this theory using phylogenetic generalized linear modeling on quantitative leaping data for 61 species collected from locomotor studies, and data from a study that recently tested Isbell's snake detection theory (Wheeler 2011). Leaping frequency and body mass had significant and positive associations with orbital convergence, while the controls (e.g. anatomy, frugivory, and nocturnality) did not. Better measures for various predation risks on primates are needed to help understand the conflicting selection pressures on orbital orientation and to resolve the evolutionary chronology of detection, evasion, and avoidance.
| 13: Schruth, D.M. (2015). "Testing the origins of primate anterior orbital convergence as a function of evasive leap landing and reduced posterior predation," American Society of Primatologists (annual meeting), American Journal of Primatology, Vol 77(S1). |

|
Humans perceive, manipulate, modify, and create numerous complex objects as a part of everyday life. Our evolved analytical perception capacities for identification of complex shapes and classification of objects enable us to use tools and communicate symbolically. The abundance of diverse tools in our environment has even been suggested as underlying the many parts of speech and prepositions that comprise our complex grammar. Recently, researchers have theorized that complex tool use may have emerged in recently terrestrialized primates who have re-purposed anatomy evolved for handling arboreal supports. Studies of complex tool use and symbolic culture are now being extended to observations of non-human Anthropoids. I hypothesize that our sophisticated symbolic communications have cognitive roots in the early Cenozoic when survival of emerging primates depended upon rapid recognition of complex branch shapes during locomotion involving precision limb landing. Leaping and arboreality substantially reduced the effectiveness of olfactory signaling, however, and may have additionally necessitated an increase in the complexity of vocal communication. Inspection of the vocalizations of leaping and brachiating primates suggests complex vocal gestures could have initially evolved as signals of visual acuity–an ancient relationship which may underlie the strong overlap between auditory and visual language systems in humans.
12: Schruth, D.M. (2015). "Precision limb landing in complex arboreal topologies as a pre-adaptation for complex symbolic communication systems," Human Behavioral Ecology Society (annual meeting). Thanks to B.Scleza for "precision"
Primates are a highly arboreal clade characterized universally by grasping appendages and hind-limb dominated climbing. Plesiadapiform fossils recently unearthed in North America implicate grasping as predating orbital convergence (and hind limb elongation) -- suggesting that leaping, a behavior hypothesized to provide the selective context for such morphology, may have been a penultimate step in the evolution of the primates of modern aspect (Euprimates). Other theories such as "visual predation" and "camouflage breaking" also predict these vision changes but additionally accommodate for the fact that the benefits of binocular parallax are limited to close range targets. I hypothesized that these close range preferences could be advantageous late in a leaping bout when urgently determining appropriate limb positioning and grasp orientation for safe landing (as destination substrate configuration may be unknown or ambiguous during take-off). Because precise knowledge of early primate leaping behavior is currently unavailable, I complied quantitative locomotor estimates for 103 species (from 42 source studies) and performed ancestral character estimation [ACE] on them to calculate a phylogenetically-weighted, average leaping frequency for the ancestral Euprimate. My analysis shows that the first true primates had repertoires with leaping composing nearly half (47%) of their locomotor bouts, strongly supporting a leaping-based theory of Euprimate morphological origins. Future origins models might benefit from a composite approach whereby elements are incorporated from multiple theories (e.g. leaping as a mechanism of snake predation avoidance).
| 11: Schruth, D.M. (2015). "Frequent Leaping Origins: unpredictable substrate orientation and position as the selective context for Euprimate visual system improvements," American Association of Physical Anthropology (annual meeting), American Journal of Physical Anthropology, Vol 156(S60). |
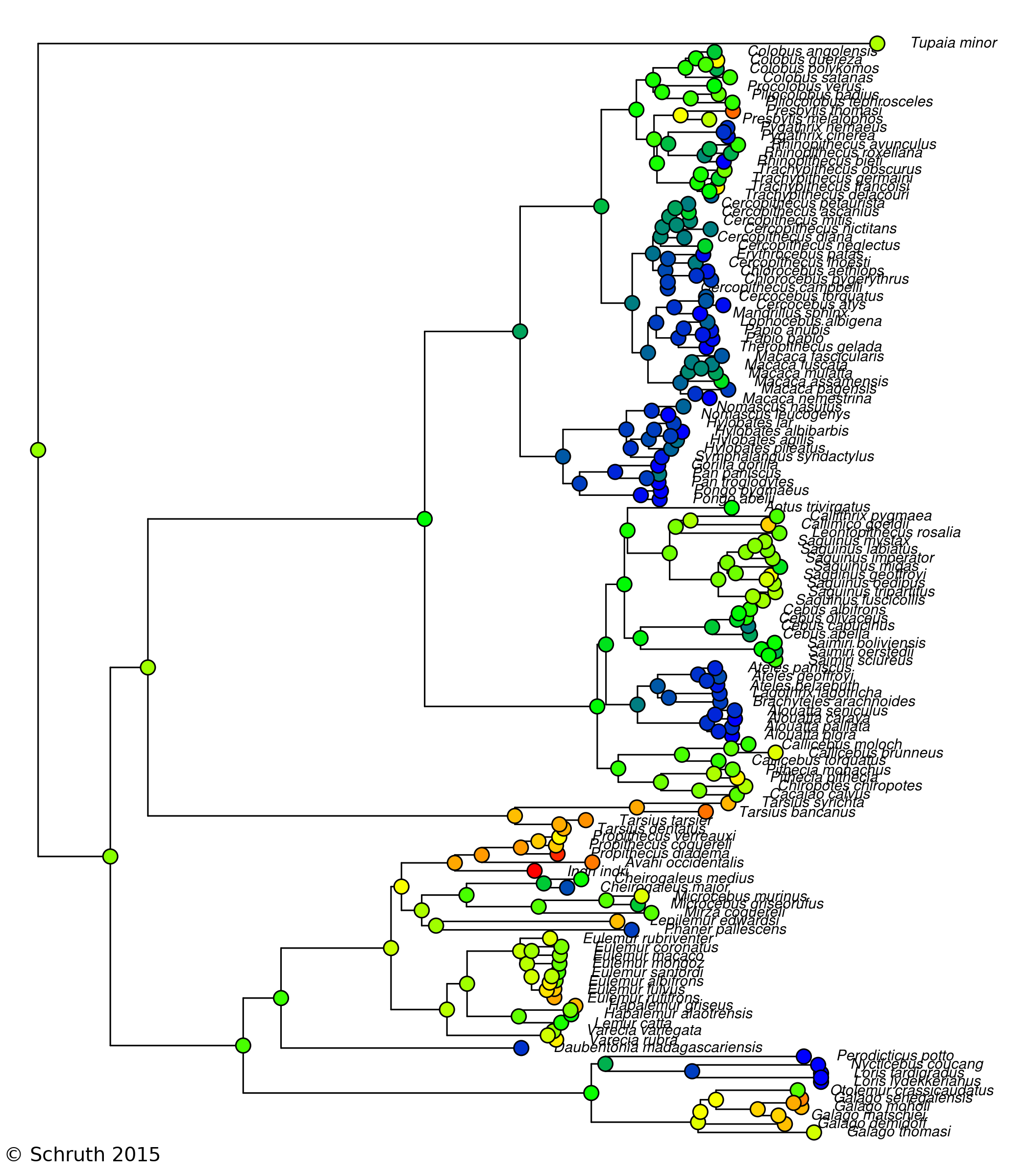
|
2014
|
10: Schruth, D.M. (2014). "Quantifying spectographic representations of primate vocalizations: the search for a unidimensional latent measure of musicality," Multivariate Data Analysis for the Social Sciences (final project). |
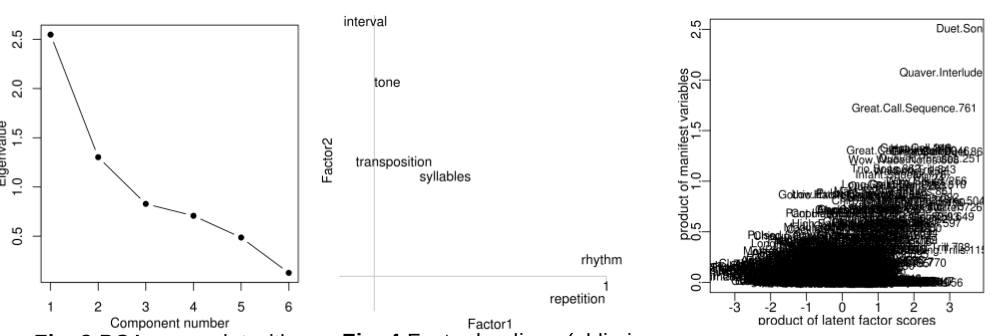
|
Theories on the origins of the unique visual and manual physiological traits of primates mostly focus on them as adaptations for grasping based locomotion through terminal arboreal substrate. The visual predation hypothesis classically invokes Lorisiforme as a counter example to the jumping based arboreal version of this class of hypothesis despite fossil and genetic evidence that frequently jumping Galagos are most similar to the ancestral primate locomotor behavioral form. Modern Galagos also differ markedly from Lorises in vocalization structure providing subtle clues to a possibly functionally coupled relationship between complex vocalization structure and precision targeted locomotor modes. Using phylogenetic contrasts and ancestral character evolution analyses we show evidence for song-like display calls (scored using vocal unit reappearance clustering) as co-evolved signals for cognitive control in timing-sensitive aerial locomotion (e.g. jumping, leaping, brachiating, & armswinging). Our analysis lends strong support for the classic arboreal hypothesis, implicates vocal display calls as important signalling mechanisms in primate behavioral evolution, and indicates that less-elaborate types of singing may have been the ancestral form for all primates.
9: Schruth, D.M., Templeton, C.T. (2014). "Singing and swinging: call structure as a function of substrate use," American Society of Primatologists Annual Meeting, American Journal of Primatology, Vol 76(S1). L.Jones-Engel suggested the three word (pre-)title
| α: Schruth, D.M. (2014). "The Origins of Song: convergent evolution, ancestral euprimates, and ecological forces that shape acoustic signal structure," Biological Anthropology Seminar Series, University of Washington. |

|
|
During the course of evolution from a generalized arboreal Archontan mammal to a more modern Euprimate, primates developed dentition for specialized frugivory, improved grasping, stereoscopic
vision, and leaping capabilities. Thumb nails, arguably the most uniquely derived trait of primates, evolved early in this process, purportedly as part of improved feeding efficiency during quadrapedal grasping locomotion on terminal branches of angiosperm flora, which were concurrently emerging in the late Cretaceous. Here I present a related yet alternative theory for the origins of primate nails centering on the similarly unique primate behavior of infant carrying. Using a simple phylogenetically controlled statistical test of data collected from only two different recent studies, I reveal a significant coevolutionary relationship (PGLM; p<.001) between infant "riding" and the number of nails (vs claws) on the hands of 50 primate species. The timing of the early emergence, exceptions (Callitrichiaes) and other possible ecological pressures (predation & canopy formation) are discussed.
8: Schruth, D.M. (2014). "Are primate nails an adaptation for infant carrying?," Anthropology S.T.A.R. Conference. University of Washington.
Primates are one of the most behaviorally diverse group of animals exhibiting a dazzling array of locomotor and communication adaptations for a vast number of differing habitats across the globe. Recently unearthed fossils such as Carpolestes (@ 85MYA), have revealed our early terminal branch friguivory adaptations but less is known about the ecology and behavior which lead to late primate morphological changes such as orbital convergence, long lower limb morphology, and increased cranial volume. Classic theory suggests that grasp-leap locomotion and/or arboreal complexity played a role in the emergence of these latter traits. Another recently theory suggests that orbital convergence evolved as a "camouflage breaking" adaptation to snake predation. Here I review the current evidence for and timing of these different theories of primate origins while comparing and contrasting current consensus with my own new vocalization and locomotion data. I weigh in supportively for a growing consensus that many of these theories and pressures each acted independently over time in truly gradual Darwinian fashion.
$: Schruth, D.M. (2014). Inferring ancient locomotor and vocal behavioral adaptations from extant primates," Anthropology S.T.A.R. Conference. University of Washington.
$: Schruth, D.M. (2014). Inferring ancient locomotor and vocal behavioral adaptations from extant primates," Anthropology S.T.A.R. Conference. University of Washington.
2013
|
δ: Schruth, D.M. (2013). "melody: statistical methods for the quantitative analysis of song spectrograms," v. 0.4.7, Comprehensive R Archive Network. |
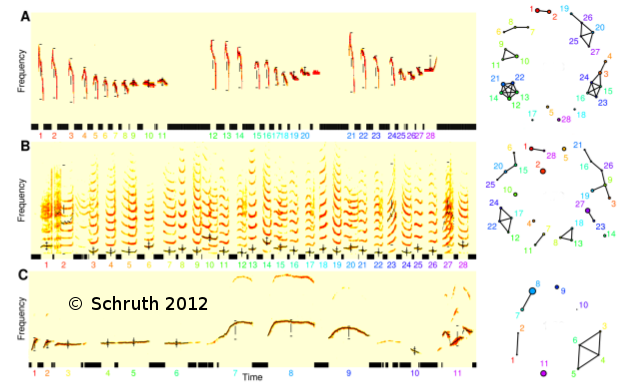
|



2012-2008
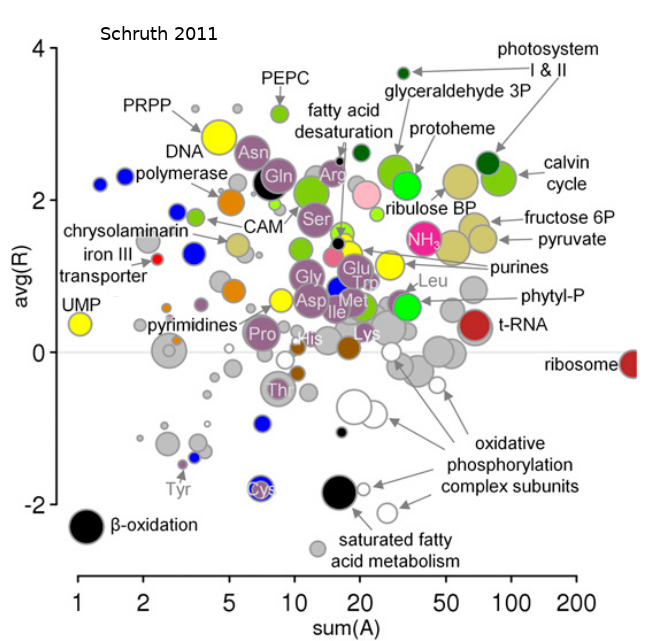
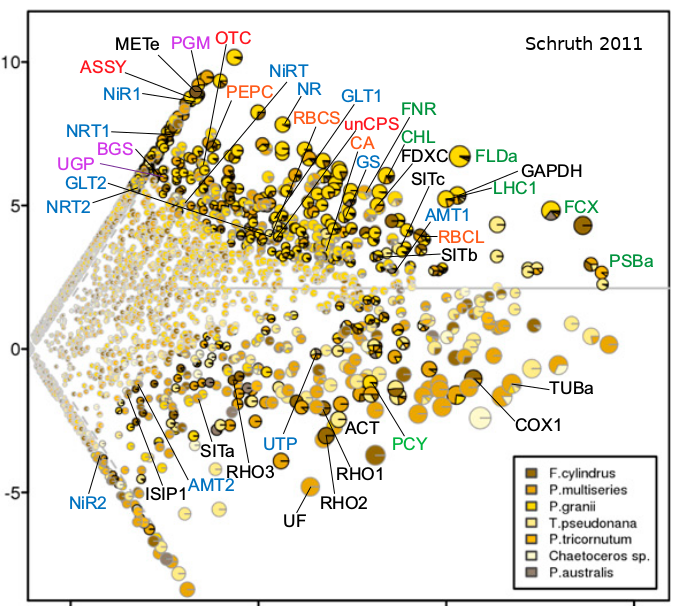

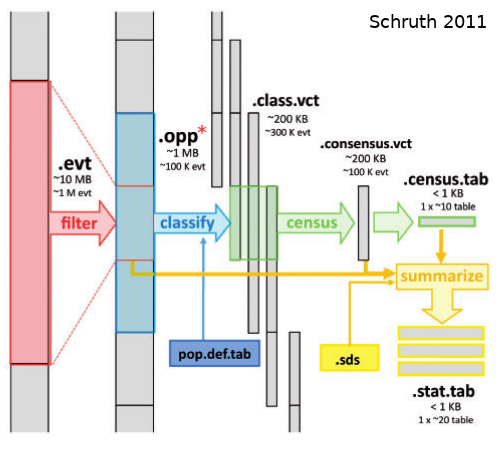
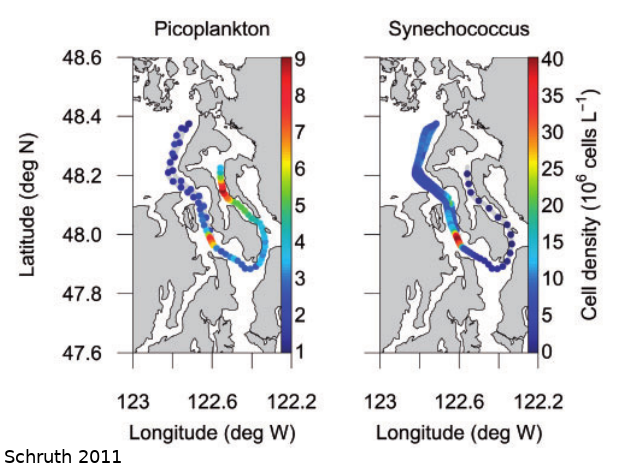
2006
The ubiquity of musical behavior in several phylogenetically distinct taxa in the animal kingdom and throughout every society in the human species is one of the greatest unsolved mysteries in biology. I have incorporated knowledge of human musical universals into a measurable working (rhythm absent) definition of melody: essentially variation and repetition of (tonal) intervals. I hypothesized that repetition and variation in melody have evolved as honest signals of underlying quality of precision and range (respectively) of distance gauging abilities. I predicted that melodic display would be positively correlated with remote targeting behaviors such as projectile locomotion (jumping, brachiation, or arm-swinging). Quantitative locomotor distributions and spectrographic vocal repertoires of primates were collected and my spectrogram analysis R-package, Gramparse, was used to split, match, and score each vocalization type. Four linear least squares regression models of (percent) projectile locomotion vs melodic score (as measured by the number of repeated interval groups per vocalization) were run: on both species level data and the corresponding phylogenetically controlled independent contrasts with and without control from additional behavioral and environmental variables. Percent projectile locomotion was statistically significant at the .01 level but failed to explain more than 20% of the variance in melodic display. This work is an important first step in both investigating possible selection pressures in a currently neglected corner music origins research and, more generally, quantifying the formerly qualitative task of assessing musicality.
|
III: Schruth, D.M. (2006). "Melodic Display as a Signal of Remote Targeting Ability," Master's Thesis. University of Washington Advisor: D.J. Holman |

|
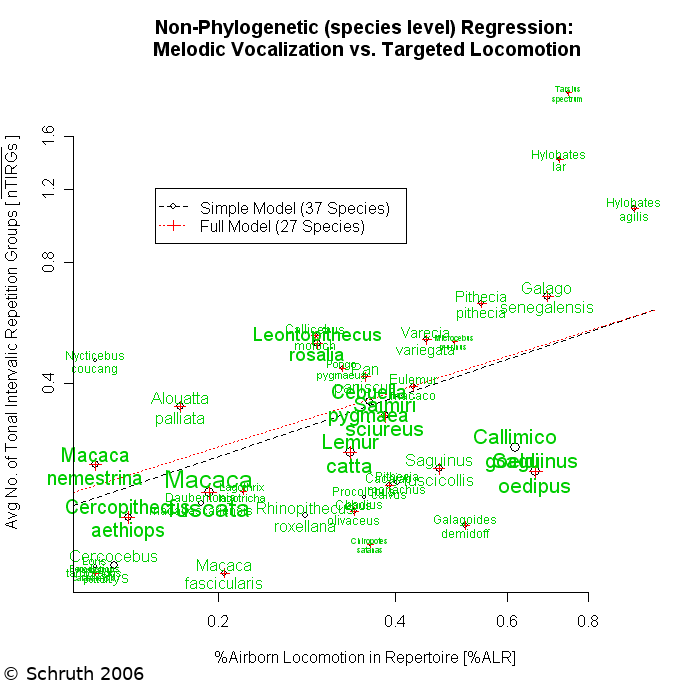
|
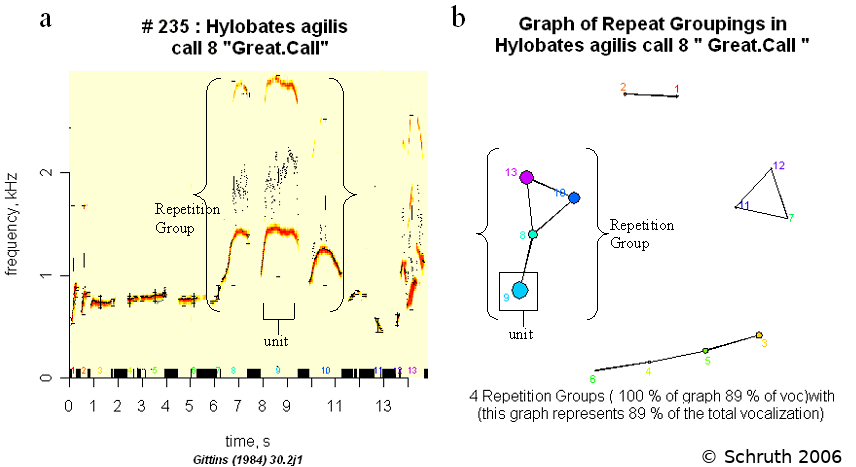
|
2005
The question of how and why music has evolved has been entertained and discussed among disciplines as disparate as Biology, Psychology, Anthropology, and Physics and has been pondered for thousands of years. Although recent literature has exploded in potential solutions to this ancient mystery, it seems that no clear answers are claiming exclusive authoritative [verity]. Problems range from the fact that there are no clear methodologies or experts as music spans so many disciplines, to the ethical, chronological, and funding problms of testing these hypotheses, especially in the case of humans. The fact that it remains such as challenge to even define "music" at all is testimony to this fact that it is a [multifarious] problem. This paper attempts to set up a general framework for explaining how music might have evolve in the human lineage. The first step is to define music in a way that is most inclusive of all human musics and yet is exclusive enough to capture some of the finer complexities which separate human musicality from the universe of normal animal communication vocalizations. The equally complex and evolutionarily convergent musics of whales, birds, and gibbons must also be addressed in this framework as the complexity and structural similarity is on par with that of most human societies. Once music is defined, and the fundamental universally occurring psychoacoustic integerial structures of music are parameterized, one can finally venture out into exploring the answers to our main questions at hand: is music adaptive? And why is it adaptive?
The first question is it adaptive, lends itself to an intuitive plea: if it is everywhere in humans and convergent in four distinct phylogenetic taxons it must have some value to its practitioners. The question with humans is a little more complicated because musical behavior might now be an exaptation, a trait that once had direct fitness benefits but that is now more of a side effect of ancestry. But still interesting and worth researching is the question of whether or not it was once adaptive and how it might have been adaptive int he environment of evolutionary adaptedness [EEA]. But as a null hypothesis, we must consider the possibility that music is not an adaptation (evolved as an adaptation at some point) and instead evolved by accident, at least in the lineage under investigation (in this case: humans). Counter-adaptationist claims must first be addressed before possible adaptations can be explored: arguments that music may have evolved out of a byproduct of some closely related adaptation (language) or other natural phenomenon (acoustic periodicity and correlated harmonic structure) are also addressed and dismissed. After null arguments are addressed and possible causes of musical adaptation are justifiably discussed. Ultimate causes of musical fitness conferring benefits are explored under the honest signaling framework (collapsing sexual and group selection into one larger phenomenon). Finally, proximate mechanisms which account for the inherent intervallic structure universally present in all four musical species are discussed. It is hypothesized that music arose in all three species as a directly encoded honest signal of spatio-temporal adaptation to rapid 3D subsistence. In the case of humans, this rapid 3D subsistence is speculated to be in the form of ballistic hunting (the high speed tool precisely projected through three dimensional space).
II: Schruth, D.M. (2005). "Ballistics and the Evolution of Human Music," Master's Thesis Prospectus. University of Washington
Advisor: D.J. Holman
Advisor: D.J. Holman


2001
Efficient Communication of Spatial Intelligence in Species Inhabiting Spatio-Temporally Demanding Environments
[Formerly: The Evolutionary Origins of Music: Communication of Spatial-Temporal Abilites in Monogamous Species]
The enormous selective pressure for spatial intelligence and an efficient way to communicate this ability provided the main stimulus for the instinctual knowledge and expression of the octave and other simple integer frequency ratios which serve as a foundation for the octave based twelve-tone scale of [western] musical theory.
Recent studies in the cognitive psychology of music have exposed a positive effect of musical training on spatio-temporal intelligence. Other studies have similarly shown that musical ability greatly improves understanding of proportional math (the math of ratios). Not surprising, then, are the separate studies showing a link between spatial intelligence and the understanding of proportions (ratios). I hope to show that the strong links between these three abilities are not merely behaviorally causal from musical activity to spatial intelligence (as has been previously shown) but evolutionarily causal from spatial intelligence to musical activity. I plan to argue that the octave (simple integer frequency ratio) based musical expression has evolved through sexual section as a [reliable] indicator of spatial-intelligence.
I: Schruth, D.M. (2001). "The Evolutionary Origins of the Simple Integer Frequency Ratios in Music," Undergraduate Honors Thesis Prospectus. University of Washington Advisor: D.J. Holman
[Formerly: The Evolutionary Origins of Music: Communication of Spatial-Temporal Abilites in Monogamous Species]
The enormous selective pressure for spatial intelligence and an efficient way to communicate this ability provided the main stimulus for the instinctual knowledge and expression of the octave and other simple integer frequency ratios which serve as a foundation for the octave based twelve-tone scale of [western] musical theory.
Recent studies in the cognitive psychology of music have exposed a positive effect of musical training on spatio-temporal intelligence. Other studies have similarly shown that musical ability greatly improves understanding of proportional math (the math of ratios). Not surprising, then, are the separate studies showing a link between spatial intelligence and the understanding of proportions (ratios). I hope to show that the strong links between these three abilities are not merely behaviorally causal from musical activity to spatial intelligence (as has been previously shown) but evolutionarily causal from spatial intelligence to musical activity. I plan to argue that the octave (simple integer frequency ratio) based musical expression has evolved through sexual section as a [reliable] indicator of spatial-intelligence.
 |
 |
 scores of pleasantness (dots) alongside SIFRs (black) |
1: Schruth, D.M., Chaloupka, V. (2000). "Consonance and Dissonance: Nature versus Culture," Undergraduate Research Symposium. V.C. suggested this title and designed the experiment; D.S. collected data, performed analysis, and wrote up the results
SOFTWARE
TEACHING
Assistant
Associate
PROFESSIONAL SERVICE/ASSOCIATIONS
Committee Member:
American Society of Primatologists: Web improvement committee
Member:
American Society of Primatologists
American Association of Physical Anthropology